三、问答题1. 剂量补偿作用
剂量补偿作用(dtosage compensation effect):所谓剂量补偿作用是使具有两份或两份以上的基因量的个体与只具有一份基因量的个体的基因表现趋于一致的遗传效应。
2. 据调查在某个偏僻的山区,有不少家庭出现连续数代的智力低下的成员,请你提出各种可能的原因,并设计一个调查研究的提纲,以便派出研究人员对该地区的这种不正常的智力低下的现象进行研究,以判断在这个人群中智力低下高频发生的原因是遗传的还是非遗传的,或者是遗传和环境相互作用的结果。
在这个人群中智力低下高频发生的原因可能是由遗传的作用,也可能是由于环境的作用,还可能是遗传和环境相互作用的结果,可通过以下调查和分析方法进行判断:
1.家系调查与系谱的绘制分析。系谱分析是将某一家族成员所得有关某一病理或生理性状发生情况的资料,按一定格式,绘制成系谱图,再对它进行研究分析,以确定所研究的特定性状在这个家庭中是否有遗传因素,及其可能的遗传方式。
2.双生子法。双生子有同卵双生与异卵双生之分。通过同卵双生子可以研究不同环境因素对表现型的影响,通过异卵双生子在相似环境中的生长、发育,可以研究不同基因型的表现型效应。通过比较同卵双生与异卵双生子某一性状发生的一致性,可以看出遗传因素和环境因素在性状发生中的比重。如在某一性状中,同卵双生与异卵双生的性状_致性无明显差异,说明此性状与遗传无关,如在某一性状中,同卵双生性状的一致性显然比异卵双生高,说明这些性状与遗传因素有关。
3.普查法(群体调查法)。如某一性状在特定的家属中发现率显著高于一般群体的发现率,说明该性状与遗传有关。
3. 酵母基因组全序列的测定业已完成,结果发现根据序列确定的相邻基因间的物理距离(千碱基对,kb)与连锁图上的相应基因间的遗传图距(厘摩,cM)不成比例,如A、B、C、D四个基因的物理距离为
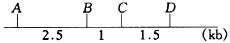
而遗传图距为
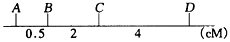
。请问这种现象是正常的,还是不正常的?可否设计更精确的遗传分析实验解决这一矛盾?如何解决?并说明理由。
1.这种现象是正常的,它是由于经典遗传学和分子遗传学关于基因概念的不同理解而引起的。按照经典遗传学对基因的概念,基因具有下列共性:
(1)基因具有染色体的主要特性:自我复制与相对稳定性,在有丝分裂和减数分裂中有规律的进行分配。
(2)基因在染色体上占有一定位置(基因座),并且是交换的最小单位,即在重组时不能再分隔的单位。
(3)基因是以一个整体进行突变的,故它又是一个突变单位。
(4)基因是一个功能单位,它控制着正在发育着的有机体的某一个或某些性状,如红花、白花等。
可以把重组单位和突变单位统称为结构单位。这样,基因既是一个结构单位,又是一个功能单位。
2.分子遗传学的发展一方面揭示了遗传密码的秘密,使基因的概念落实到具体的物质上,获得了具体的内容。另一方面,在精密的微生物遗传分析中查明,基因并不是不可分割的最小遗传单位,而是远为复杂得多的遗传和变异单位。例如,在一个基因的区域内,仍然可以划分出若干个起作用的小单位。按照现代遗传学的概念,重组、突变、功能这三个单位应该分别是:
(1)突变子(muton):它是性状突变时,产生突变的最小单位。一个突变子可以小到只是一个核苷酸对。
(2)重组子(recon):在发生性状的重组时,可交换的最小单位称为重组子。据微生物重组的精细研究证明,一个交换子只包含一对核苷酸。
(3)顺反子(cistron):这一术语表示一个起作用的单位,基本上符合通常指的基因或略小的单位。一个顺反子所包括的一段DNA与一个多肽链的合成相对应。平均大小为500~1500个核苷酸。
由此可知,过去作为结构单位的基因,实际上包含大量的突变子或重组子。以往认为基因是最小的结构单位,现已不能成立了。然而关于基因是一个功能单位的概念仍然是正确的。
3.物理距离为分子遗传学关于基因概念的范畴。基因内部可能含有内含子,而遗传图距是经典遗传学关于基因概念的范畴,所以两者不一致是正常的。为了解决这一矛盾,可设计以下更精细的遗传分析实验。
(1)互补作用。假定有两个独立起源的隐性突变,它们具有类似的表型。如何判断它们是属于同一个基因的突变,还是分别属于两个基因的突变?即如何测知它们是等位基因?为此,需要建立一个双突变杂合二倍体,然后测定这两个突变间有无互补作用(complementation)。如有互补作用,个体应表现为野生型;如无互补作用,则个体表现为突变型。这是因为如果这两个突变型来自一个基因,则两条同源染色体都只能产生突变的mRNA,其个体表现型应是野生型。
这种根据功能确定等位基因的测验称为互补测验或顺反测验。原来,杂合的双突变体有两种不同的排列形式:顺式排列和反式排列。顺式排列是指两个突变基因座在同一条染色体上,反式排列是两个突变基因座在不同的染色体上。顺反测验就是根据顺式表现型和反式表现型来确定两个突变体是否属于一个基因或顺反子。实际上,顺式排列只是作为对照,一般并不进行测试,因为它的表现型永远是野生型。测验实质上是反式测验。如反式排列表现为野生型,说明这两个突变分别属于两个基因基因座,即非等位基因;如表现为突变型,则说明两个突变属于同一个基因的不同基因座,即等位基因。
(2)基因的微细结构。20世纪50年代的生化技术还无法进行DNA的序列测定,因此本泽尔利用经典的噬菌体突变和重组技术.,对T4噬菌体rⅡ区基因的微细结构进行了详细分析,为研究基因的微细结构提供了范例。在噬菌体染色体的三个不同部位上有三个不同的r突变型:rⅠ、rⅡ和rⅢ。研究最清楚的是rⅡ突变型。野生型T4噬菌体侵染大肠杆菌B株和K12(λ)株(带有整合到大肠杆菌染色体上的λ噬菌体),经6~10小时形成小而边缘模糊的噬菌斑;而rⅡ突变型T4噬菌体,在侵染大肠杆菌20分钟后,即形成大而边缘清楚的噬菌斑。但是rⅡ突变型只能在B株上生长,不能在K12(λ)上生长。就是利用这个特点,让不同的两个rⅡ突变型杂交,然后在K12(λ)株上用选择方法把重组体r+筛选出来,从而计算出这两个r+突变基因座间的重组频率。具体作法如下:
用大量的rⅡ突变体对大肠杆菌B株成对进行双重感染(double infection),这里强调要同时,不能一先一后,否则出现排斥现象。超数感染的噬菌体迅速被宿主的DNA酶所破坏,不能参加重组。形成噬菌斑后,收集溶菌液(内含子代噬菌体),把它接种到B株上,计算溶菌液中的总噬菌体数,因为两种rⅡ突变体(rx、ry)、重组体(r+r+)和(rxry)都可以在B株生长。同时把溶菌液也接种到K12(λ)株上,计算野生型重组体r+r+数目,因为只有r+r+可以生长,而其余三种基因型不能生长(包括重组体rxry),所以在计算重组体数目时,总要乘以2,就是因为rxry虽然是预期的,但不能检出。
重组值的计算方法:
重组值=[2×(r+r+噬菌体数)/总噬菌体数]×100%
=[2×(在K12(λ)株上生长的噬菌斑数)/在B株上生长的噬菌斑数]×100%
用本法可以检出小到0.001%,即十万分之一的重组值。根据大量的二点杂交法所得的重组值,去掉%,即为两个突变座位间的距离。利用大量rⅡ区内二点杂交的结果,可绘制出rⅡ区基因座间微细的遗传学图。这同二倍体通过一系列二点杂交法绘制出的连锁图是相同的。
必须指出,本泽尔所用的rⅡ突变型可分成A组和B组两类,只有当一个A组和一个B组的突变体混合感染K12(λ)时,才发生溶菌现象,即互补现象。而用两个A组突变体或两个B组突变体则没有溶菌现象,即不发生互补作用。
4. 遗传学的发展是与模式生物的应用分不开的,如孟德尔的豌豆试验、摩尔根的果蝇试验。试举出另外二种模式生物,说明它们在推动遗传学发展中的作用。遗传学的创新与试验材料的创新有什么关系?
细菌和病毒也是遗传学中的模式生物,它们在遗传研究中的优越性及在推动遗传学发展中的作用可以归纳为:
(1)繁殖世代所需时间短。每个世代以分钟或小时计算。如病毒每1小时可繁殖成百,大肠杆菌每20分钟可繁殖一代。
(2)易于管理和进行化学分析。用一支试管可以储存数以百万计的细菌和病毒,操作管理方便,可大量节省空间和培养工作所需的人力、物力、财力。在基因作用的研究上常需要对代谢产物或基因本身进行化学分析,而细菌代谢旺盛,繁殖又快,可在短期累积大量产物,为化学分析提供了条件。
(3)遗传物质比较简单。细菌和病毒的遗传物质包括一个位于细胞质内裸露的DNA分子(病毒有时是RNA分子),它与真核生物不同,没有组蛋白及其他蛋白质的结合。因为缺乏适当的名词,现仍称之为染色体。这类染色体更适宜用作基因结构和功能的研究以及基因工程的操作。关于基因精细结构的研究,首先需要得到一个基因内部大量不同位点(site)的突变型,然后通过重组决定它们的顺序。由于这些突变可能以突变子(muton)为单位,它们之间距离很近,所以必须观察极大量的子代,才能看到少数的重组体。只有用细菌和病毒作材料才能满足这个要求。
(4)便于研究基因的突变。细菌和病毒属于单倍体,所有突变都能立即表现出来,不像真核二倍体生物那样,有显性掩盖隐性的问题。此外,基因的突变频率很低,必须有大量的个体才能看到一个突变型。如一个培养皿有几百个菌落,而突变率一般小于10-5,因此,至少需要上百个培养皿才能看到一个突变型。如果观察的是抗四环素突变型,只要在培养基上加入四环素,则敏感的野生型细菌不能形成菌落,只有发生抗性突变的细菌才能长出菌落。因此,用此法不难从若干亿个个体中选出个别突变。
(5)便于研究基因的作用。细菌可以生活在基本培养基上,易于获得营养缺陷型,也易于测知各种营养缺陷型所需要的物质,是研究基因作用的好材料。
(6)可用作研究高等生物的简单模型。高等生物体内机制复杂,难于着手进行复杂的遗传学研究,如基因表达的调控问题等。而细菌和病毒的结构简单,较易分析研究,可以从微生物的研究中得到模型,以便从中获得启发,从而开展高等生物的遗传研究。
正因为细菌和病毒具有以上优越性,1944年Avery等人用肺炎链球菌证实了遗传物质是DNA。1952年Hershey等用噬菌体侵染细菌的实验进一步证实了基因就是DNA,极大地推动了分子遗传学的发展。1965年法国科学家Jacob和Monod用大肠杆菌为试验材料研究乳糖代谢的调控,提出并证实了操纵子作用调节细菌代谢的分子机制,他们的这一学说对分子遗传学的发展起了极其重要的指导作用。可见试验材料创新促进了遗传学的发展。
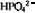 及
及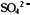 和一种碳源,如葡萄糖、甘油或乳酸盐等。
和一种碳源,如葡萄糖、甘油或乳酸盐等。

 深色:已答题 浅色:未答题
深色:已答题 浅色:未答题